Study on the development from embryo to adult stages in the Naru eagle ray, Aetobatus narutobiei, in the western Seto Inland Sea
Akiko HIJIYA, Aya SHIMAMOTO, Saki MINPO, Hiromi YAMASAKI, Norio SHIMIZU, Akihito WANISHI, Yoichi SAKAI and Hiroaki HASHIMOTO
Note: Published in Bulletin of the Hiroshima University Museum 2021, the original English abstract is attached below. The following parts of this article are translated and summarized from the Japanese content of the publication.
At the end, you can find numbers about the Naru eagle ray census conducted by the government.
Abstract
Since the beginning of the 21st century, the Naru eagle ray (Aetobatus narutobiei) has been found to be distributed in large numbers in the western sea regions of Japan. Moreover, it has been exterminated as it feeds on beneficial shellfish. It is an ovoviviparous ray, and ecological studies on this species have been carried out around the Japanese coast; however, the developmental stages in the life of the Naru eagle ray have not been reported yet.
Based on our research in the western Seto Inland Sea, Japan, from 2004 to 2012 on a total of 368 embryos (uterus larvae) and 985 free-swimming individuals, we identified the developmental stages of this ray according to those of marine teleosts (stages: egg, larva, juvenile, young, immature, and mature).
As a result, the Naru eagle ray embryo was found to develop from an egg and pass through the juvenile to the young stage in the uterus of the female parent, where it utilizes the yolk and ingests the uterine milk through the trophonemata. Before birth, the uterus larva completes its body form as a species.
This species exhibits sexual dimorphism, where females grow slower, larger, and live longer than males. It could be considered that sexual dimorphism along with the modes of development and maintenance of the uterus larva are evolutionarily conserved mechanisms.
Background
The Narutobiei (Aetobatus narutobiei), a species of cartilaginous fish in the family Myliobatidae, was first recorded in Japan in 1989. Since the early 2000s, it has appeared in large numbers in areas like the Ariake Sea and western Seto Inland Sea, where it has damaged commercially important bivalves such as Ruditapes philippinarum (clams) and Atrina pectinata (pen shells), prompting control efforts.
However, as the species progressively became viewed as potential food, some actors advocated treating it as a harvestable resource rather than a pest and it slowly became a “capture target” rather than “eradication” target. In 2017, Japan’s Ministry of the Environment and Fisheries Agency designated the species as endangered.

Introduction
Like other rays in the order Myliobatiformes, the Narutobiei exhibits aplacental viviparity, initially relying on yolk and later on “uterine milk” (a nutrient-rich secretion) for embryonic development. Although biological studies have explored its reproduction, growth, diet, and distribution, especially in the Ariake and Seto regions, research focused specifically on developmental stages is scarce, with only a few studies addressing in-utero development. Development in fish is understood as a qualitative, non-repetitive, stage-based process distinct from growth, which is quantitative.
The authors analyzed specimens from 2004–2012 in western Japan using morphometric and ecomorphological approaches to map the full developmental trajectory of A. narutobiei, from embryo to adult. Their work also draws on marine fish developmental stage classifications (egg → larva → juvenile → young → immature → mature) and compares A. narutobiei with related species to highlight its unique features.

Ruditapes philipinarum & local Onoda clams

Atrina pectinate
Research method
Between 2004 and 2012, researchers collected specimens of the Narutobiei (Aetobatus narutobiei) from the coastal waters around Kariyagawa Port in Sanyo-Onoda City, Yamaguchi Prefecture, as part of an annual ray removal program conducted jointly with the local fisheries cooperative. This program began in 2003 in response to significant predation by the rays on the locally prized “Onoda Asari” clams. The rays were captured using drift gillnets intended for pomfret fishing.
Specimens, including intrauterine larvae expelled due to capture shock, were collected primarily from spring through autumn, typically during monthly onboard surveys. Researchers measured disc width (DW) with tape measures and recorded sex based on clasper presence in males. Intrauterine larvae—ranging from early stages with external yolk sacs to those near birth—were weighed with high precision scales, and yolk sac and intestine weights were also measured when possible. Some larvae exhibited distended abdomens due to dark green intestinal contents, which solidified in formalin for further analysis.
Developmental features such as caudal spines were categorized by formation stage, and male reproductive maturity was assessed via clasper length and calcification status. To evaluate growth rates, the study used Bertalanffy growth models (from Shimamoto et al., 2019) to compare disc width across age groups. Some specimens were examined in laboratories, while others were studied onsite, and larger individuals were weighed using body-weight subtraction methods.
Results
In the study’s results section, researchers made key observations regarding the reproductive and developmental characteristics of Aetobatus narutobiei. On May 30, 2006, mature female rays were observed releasing multiple yellow eggs (2–3 cm in diameter) encased in a translucent, gelatinous membrane from their cloacas. Later, on September 11, 2006, ovarian inspection revealed several mature oocytes (~2 cm) and dozens of smaller immature eggs (~a few mm), indicating ongoing reproductive activity.
Intrauterine larvae collected on June 20, 2006, with disc widths (DW) under 5 cm, were soft, semi-transparent, and milky-white, connected to an external yolk sac via a central yolk stalk. At this early developmental stage (around 3 cm DW), sex could already be distinguished based on the presence or absence of claspers. Analysis of larval DW distributions from 2004 and 2005 showed distinct size modes at 14–18 cm, 22–24 cm, and 30–32 cm, suggesting synchronized growth within cohorts collected between June and August. No significant differences in size or sex ratio were found across samples (t-test and chi-square test, p > 0.1 and p > 0.5, respectively).
To explore growth patterns, the researchers analyzed the relationship between DW and body weight (BW) across both sexes from larval to adult stages using data from 2005–2012. They observed that few males exceeded 100 cm DW, and regression curves showed sex-based growth trends. Using a von Bertalanffy growth model from Shimamoto et al. (2019), they calculated age-specific DW and annual growth rates, visualizing these data in graphical form to illustrate the species’ growth trajectory over time.
Individuals estimation
The estimated number of migrating Aetobatus narutobiei individuals based on the DOIRAP method was approximately 400,000 to 500,000 during fiscal years 2008 to 2010. However, from fiscal year 2011 onward, the number declined, and since fiscal year 2012, it has generally remained between 100,000 and 200,000 individuals (see Figures below).
Note that 1万 stands for 10’000 individuals, 20万 for 200’000 etc.
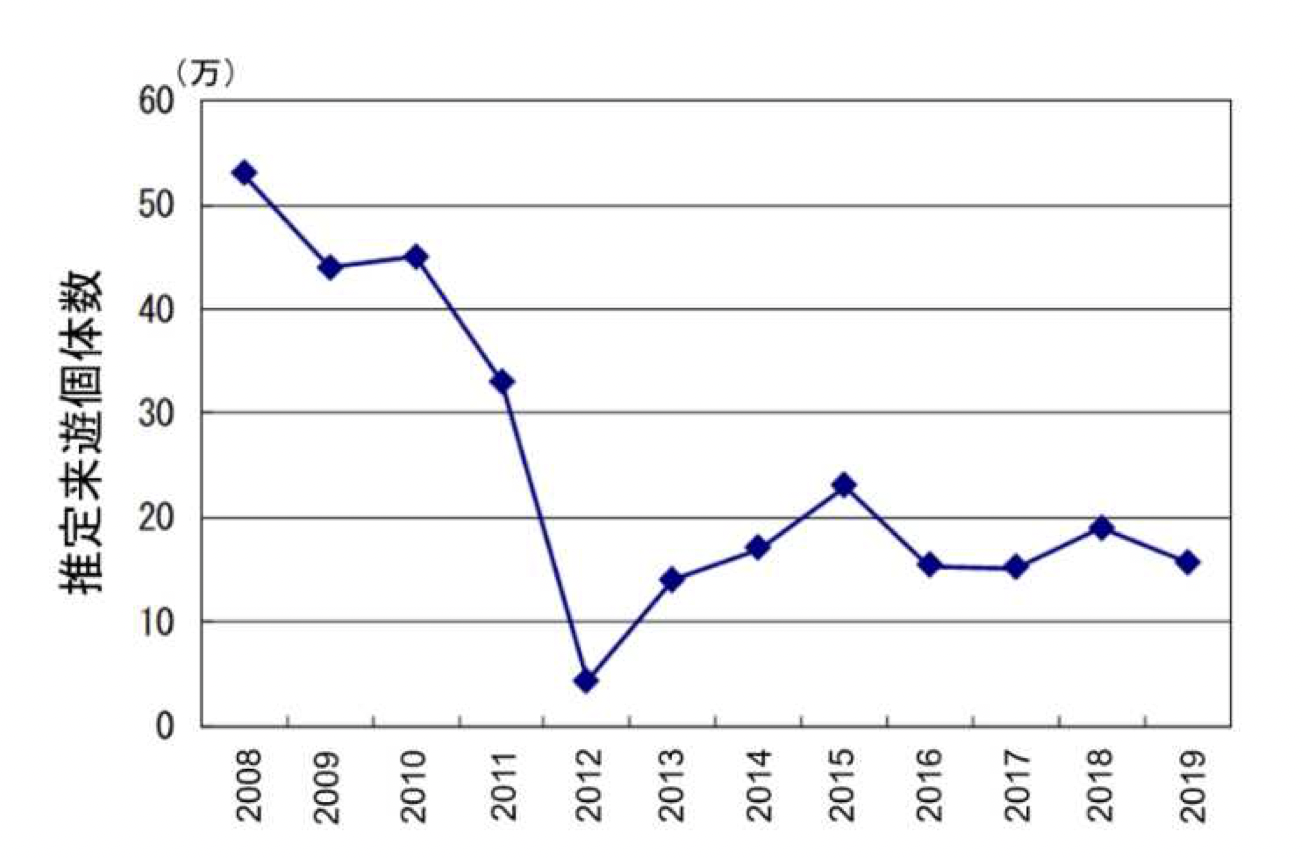
11.1-2 Changes in estimated migratory population of eagle rays over time (DOIRAP method)